



Lebensraumnutzung des Rehwilds
 Zoombild vorhanden
Zoombild vorhanden
© Soru Epotok-Fotolia.com
Das jahreszeitliche Verhalten unserer Rehe hängt von vielen Faktoren ab und ist für fast jedes Revier individuell ausgeprägt. Das Reh kann sich sehr gut an die Gegebenheiten anpassen. Diese können natürlichen Ursprungs sein wie zum Beispiel durch Witterung, Nahrungsqualität und -quantität, oder menschlich bedingt, zum Beispiel durch Freizeitverhalten oder die Jagd.
Zusammenfassung des Artikels
Rehe wandern innerhalb ihres Streifgebiets. Je ausgeprägter die Witterungsunterschiede im Jahresverlauf sind, das heißt je extremer der Unterschied zwischen Sommer und Winter ausfällt, zum Beispiel in den Alpen, desto wahrscheinlicher wandern die Rehe zwischen Sommer- und Wintereinständen.
Der halbjährliche Wechsel hat einige Vorteile, er führt zu einer Herabsetzung der Populationsdichte – und entlastet somit die Nahrungsquellen. Für Beutegreifer wird eine Bejagung des Rehwildes damit ebenfalls erschwert.
Der Wechsel von Böcken und Geißen in die Sommereinstände findet weitestgehend zeitgleich statt. Im Herbst hingegen nehmen die Geißen als erste die Wintereinstände an.
Theoretische Einführung in die Raumnutzung von Wildtieren
Die Untersuchung des Raumnutzungsverhaltens von Wildarten stellt eine wichtige Grundlage für ein innovatives Wildtiermanagement dar. So ist es wichtig zu verstehen, wie Wildtiere im Raum verteilt sind und wie sich ihre Bewegungsmuster in Abhängigkeit von bestimmten Faktoren verändern.
Räumlich wird im klassischen Sinne zwischen vier Maßstabsebenen der Lebensraumnutzung unterschieden (Johnson 1980):
- Die erste Ebene bestimmt das gesamte Verbreitungsgebiet einer Art (geographic range).
- Die zweite Ebene betrifft die Wahl eines Streifgebiets innerhalb der Landschaft (landscape scale).
- Die dritte Ebene betrifft die Raumnutzung innerhalb eines Streifgebiets (homerange scale).
- Die vierte Ebene beschreibt die Wahl einzelner, sehr spezifischer Komponenten innerhalb des Streifgebiets, wie zum Beispiel Äsungs- oder Setzplätze.
Ein Wildtier muss zwischen der Aufnahme hochwertiger Nahrung, Gefahren durch Raubtiere oder Parasiten und einem optimalen Sozialverhalten abwägen. Auf allen Ebenen dominieren verschiedene Faktoren. Wildtiere versuchen diese zu optimieren, um die besten Bedingungen für die eigene Vermehrung zu erhalten (Senft et al. 1987, Wiens 1989). Letztendlich ist das Ziel eines jeden Wildtieres, seine Fitness zu maximieren. Hier ist jedoch nicht die Sportlichkeit gemeint, sondern die Fitness im biologischen Sinn: englisch "fitness" heißt auf deutsch "Tauglichkeit". Darunter verstehen wir so oft wie möglich die eigenen Gene an zukünftige Generationen weitergeben zu können.
Raumnutzung von Rehen
Das Rehwild wechselt im Jahresverlauf zwischen zwei Phasen, zu welchen sich der Organismus und das Verhalten ändern: die Fortpflanzungsphase im Frühjahr, wenn das Rehwild überwiegend einzeln oder, je nach Habitat, in kleinen Gruppen vorkommt. Während dieser Zeit sind die Böcke in der Regel territorial, was das Rehwild von allen anderen einheimischen Schalenwildarten unterscheidet.
Im Herbst und Winter (Winterphase) schließt sich das Rehwild dagegen zu Gruppen verschiedener Größe zusammen (Hewison et al. 1998). Die genannten Eigenschaften zu den Nahrungsbedürfnissen und dem Sozialverhalten bestimmen somit das Raumnutzungsverhalten von Rehen.
Diese Anpassungsfähigkeit des Rehwildes zeigt sich unter anderem in der Unterschiedlichkeit der Bewegungsmuster in verschiedenen Lebensräumen (Peters 2015). Viele dieser Bewegungsmuster wurden erst durch die GPS Satellitentelemetrie erforscht.
Untersuchung der Raumnutzung mittels Satellitentelemetrie
Das Eurodeer-Projekt besteht aus mehr als 30 Forschungsgruppen in ganz Europa. Den Kern des Projektes bildet eine gemeinsame Datenbank, welche sowohl telemetrische Positionsdaten also auch Informationen zu den Individuen, Populationen und den Lebensräumen beinhaltet. So können verschiedenste Fragestellungen über Regionen mit unterschiedlichen Lebensräumen und deren Einfluss auf die Raumnutzung untersucht werden.
Migration: saisonale Wanderungen zwischen Sommer- und Winterstreifgebiet
Interessanterweise sind auch bei unseren Schalenwildarten solche Wanderungen zwischen saisonalen Streifgebieten nichts Ungewöhnliches. Die saisonale Migration stellt im Grunde eine Form der Lebensraumnutzung innerhalb des Streifgebiets (Raumnutzung der zweiten Ebene) eines Individuums dar.
So nutzt es innerhalb seines Streifgebiets verschiedene, räumlich und zeitlich getrennte Sommer- und Wintereinstände. Insbesondere eine hohe Saisonalität des Lebensraums, wie zum Beispiel in alpinen Gebieten, bringt periodische Engpässe mit sich. Es fällt Schalenwildarten in solchen topographisch und klimatisch anspruchsvollen Lebensräumen oftmals schwer, ihre Nahrungsbedürfnisse in der kalten Jahreszeit zu stillen. Zunehmende Schneehöhen können den Zugang zur Nahrung verhindern und auch die Fortbewegung und die Thermoregulation von Schalenwild beeinträchtigen (Cagnacci et al. 2011). Je nachdem wie stark diese Engpässe ausgeprägt sind und wie hoch die Vorhersagbarkeit der Saisonalität von Jahr zu Jahr ist, treten saisonale Wanderungen und verschiedene Übergangsformen auf (Mueller et al. 2011).
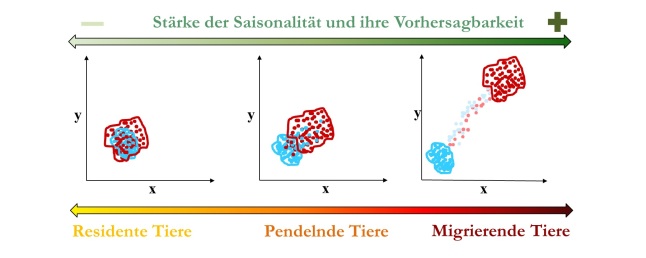
Abbildung oben: Zusammenhang zwischen Saisonalität eines Lebensraumes und der Vorhersagbarkeit der Saisonaliät zwischen den Jahren und dem Gradienten von Bewegungsmustern innerhalb einer Population. Die roten Punkte und Linien stehen für Positionsdaten und Streifgebiete im Sommer und die blauen Punkte und Linien stehen für den Winter. Die Punkte sind entlang von xy-Koordinaten dargestellt. Mit zunehmender Saisonalität und Vorhersagbarkeit dieser nimmt der Anteil von migrierenden oder wandernden Tieren innerhalb der Population in der Regel zu.
Es wird angenommen, dass die Schwierigkteiten der langen Wanderbewegungen der Tiere vom Wintereinstand zum Sommereinstand durch eine höhere und länger andauernde Äsungsqualität im Sommereinstandsgebiet kompensiert werden (forage maturation hypothesis; Hebblewhite et al. 2008).
Weitere Vorteile, welche sich wandernde gegenüber standorttreuen Tieren verschaffen könnten, sind geringere Vorkommen von Raubtieren (Fryxell and Sinclair 1988) und Parasiten (Folstad et al. 1991).
Die genannten Faktoren stehen in vielfältigen Wechselwirkungen mit der Populationsdichte (Kaitala et al. 1993, Mysterud et al. 2011) und der klimatischen Veränderunge des Lebensraums (Jonzén et al. 2011). Das Wanderverhalten und deren beeinflussenden Faktoren sind besonders interessante Aspekte der Raumnutzung von Wildtieren.
Wanderungen von Rehen im Jahresverlauf
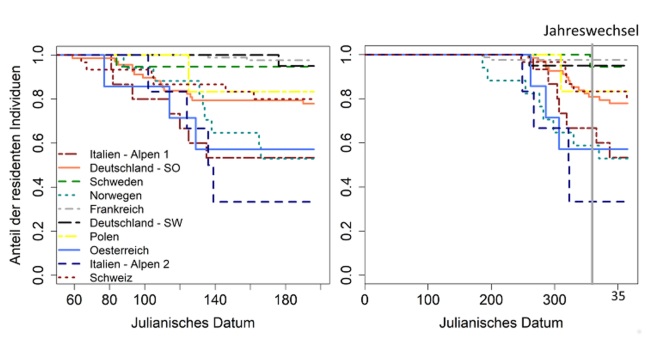
Abbildung oben: Die Abbildungen zeigen den Anteil der migrierenden GPS-telemetrierten Tiere in jedem Untersuchungsgebiet. Zum Beispiel sieht man in der linken Grafik, dass am Anfang des Jahres noch alle Rehe in ihrem Winterstreifgebiet sind. Nach und nach wandern Tiere der einzelnen Populationen ab, um in ihre Sommerstreifgebiete zu ziehen. In keiner Population wandern alle Tiere und auch in der Italienischen Population mit den meisten migrierenden Rehen sind ca. 30 % der Rehe resident. Neben dem Anteil der migrierenden Tiere zeigen die Grafiken also auch die Zeitpunkte der Migration im Jahresverlauf. Die Abbildung auf der linken Seite zeigt die Migrationszeitpunkte der Rehe im Frühjahr und die rechte Grafik zeigt den Migrationszeitpunkt im Herbst.
Die Böcke hingegen verweilen in den Sommerstreifgebieten so lange wie möglich, möglicherweise in der Hoffnung, dort auch überwintern zu können und ihre Territorien auch im nächsten Jahr zu besetzen. So kann die höhere Standorttreue der Böcke gegenüber den Ricken mit ihrer Neigung zum eigenen Revier erklärt werden (Hewison et al. 1998, Cagnacci et al. 2011).
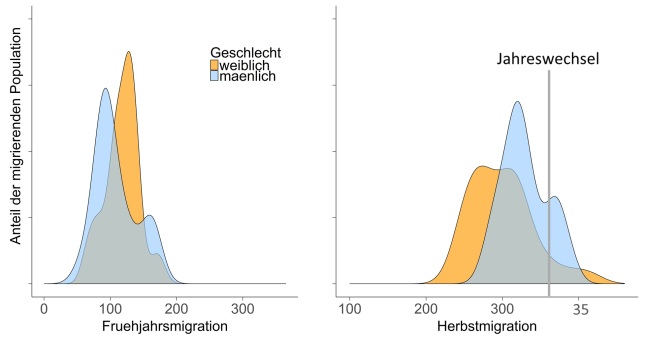
Abbildung oben: Zeitpunkt der Wanderung im Jahresverlauf, unterschieden nach Geschlechtern. Die Abbildung auf der linken Seite zeigt den Zeitpunkt der Wanderung im Frühjahr und die rechte Grafik zeigt den Zeitpunkt im Herbst.
Literaturverzeichnis
- Bischof, R., L. E. Loe, E. L. Meisingset, B. Zimmermann, B. Van Moorter, and A. Mysterud. 2012. A Migratory Northern Ungulate in the Pursuit of Spring: Jumping or Surfing the Green Wave? American Naturalist 180:407-424.
- Cagnacci, F., S. Focardi, M. Heurich, A. Stache, A. J. M. Hewison, N. Morellet, P. Kjellander, J. D. C. Linnell, A. Mysterud, M. Neteler, L. Delucchi, F. Ossi, and F. Urbano. 2011. Partial migration in roe deer: migratory and resident tactics are end points of a behavioural gradient determined by ecological factors. Oikos 120:1790-1802.
- Duncan, P., H. Tixier, R. R. Hofman, and M. Lechner-Doll. 1998. Feeding strategies and the physiology of digestion in roe deer. Pages 91-116 in R. Andersen, P. Duncan, and J. Linnell, editors. The European roe deer: the biology of success. Scandinavian University Press, Oslo.
- Folstad, I., A. C. Nilssen, O. Halvorsen, and J. Andersen. 1991. Parasite avoidance: the cause of post-calving migrations in Rangifer? Canadian Journal of Zoology 69:2423-2429.
- Fryxell, J., and A. R. E. Sinclair. 1988. Causes and consequences of migration by large herbivores.
- Fryxell, J. M., J. Greever, and A. R. E. Sinclair. 1988. Why are ungulate so abundant. American Naturalist 131:781-798.
- Hebblewhite, M., E. H. Merrill, and G. McDermid. 2008. A mutli-scale test of the forage maturation hypothesis for a partially migratory montane elk population. Ecological Monographs 78:141-166.
- Hewison, A. J. M., J.-P. Vincent, and D. Reby. 1998. Social organisation of European roe deer.
- Hofmann, R. R. 1989. Evolutionary Steps of Ecophysiological Adaptation and Diversification of Ruminants - a Comparative View of Their Digestive-System. Oecologia 78:443-457.
- Johnson, D. H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65-71.
- Jonzén, N., E. Knudsen, R. Holt, and B. Sæther. 2011. Uncertainty and predictability: the niches of migrants and nomads.in E. J. Milner-Gulland, J. Fryxell, and A. R. E. Sinclair, editors. Animal Migration: A Synthesis. Oxford University Press, Oxford.
- Kaitala, A., V. Kaitala, and P. Lundberg. 1993. A Theory of Partial Migration. American Naturalist 142:59-81.
- Mueller, T., K. A. Olson, G. Dressler, P. Leimgruber, T. K. Fuller, C. Nicolson, A. J. Novaro, M. J. Bolgeri, D. Wattles, S. DeStefano, J. M. Calabrese, and W. F. Fagan. 2011. How landscape dynamics link individual to population-level movement patterns: a multispecies comparison of ungulate relocation data. Global Ecology and Biogeography 20:683-694.
- Mysterud, A., L. E. Loe, B. Zimmerman, R. Bischof, V. Veiberg, and E. Meisingset. 2011. Partial migration in expanding red deer populations at northern latitudes – a role for density dependence? Oikos 120:1817-1825.
- Peters, W. 2015. Run to the Hills - Partial Migration in Large Herbivores. University of Montana
- Rivrud, I. M., R. Bischof, E. L. Meisingset, B. Zimmermann, L. E. Loe, and A. Mysterud. 2016. Leave before it's too late: anthropogenic and environmental triggers of autumn migration in a hunted ungulate population. Ecology 97:1058-1068.
- Senft, R. L., M. B. Coughenour, D. W. Bailey, L. R. Rittenhouse, O. E. Sala, and D. M. Swift. 1987. Large herbivore foraging and ecological hierarchies. Bioscience 37:789-795 & 798-799.
- Wiens, J. A. 1989. Spatial Scaling in Ecology. Functional Ecology 3:385-397.